
北京家恩德运医院遗传中心再发两篇SCI:从胚胎致死到复发性流产的遗传解码
2026年,北京家恩德运医院医学遗传中心在生殖遗传领域连续取得两项重要研究成果。3月,研究团队在《Frontiers in Genetics》(JCR二区,IF=3.2)发表论文,首次报道PADI6基因"一截一延"复合杂合移码突变导致着床前胚胎致死,从分子层面揭示了反复IVF失败、胚胎卵裂期发育阻滞的遗传根源;6月,团队又在《Frontiers in Endocrinology》(JCR一区,IF=5.7)发表论文,首次在家系层面证实BUB1B基因杂合变异与不明原因复发性流产的因果关联,并阐明"父系传递、女性发病"的性别特异性外显机制。两项研究分别聚焦胚胎发育的"着床前"与"着床后"两个关键阶段,共同构筑起女性生殖障碍遗传诊断的完整防线,为从"反复失败"走向"精准诊疗"提供了坚实的分子依据。
研究背景:女性生殖失败的"着床前—着床后"遗传谜题
女性生殖障碍的临床表现跨度极大:部分患者反复取卵、反复受精,胚胎却在着床前即发生发育阻滞,始终无法获得可移植胚胎;另一些患者虽能成功着床,却反复经历自然流产。前者被称为着床前胚胎致死(Preimplantation Embryonic Lethality,PREMBL),后者则为复发性流产(Recurrent Pregnancy Loss,RPL)。两者均给家庭带来沉重的身心负担,且约半数患者经全面检查仍无法明确病因,长期陷入"诊断漂流"的困境。
越来越多的证据表明,这两类看似不同的生殖失败,其根源往往可追溯至特定的基因变异。PADI6基因编码的蛋白是维持卵母细胞细胞质晶格(cytoplasmic lattices)结构完整性的关键组分,对母源蛋白储存、母源-合子基因组转换(MZT)及早期胚胎发育不可或缺,其突变可导致PREMBL2型(OMIM #617234)。BUB1B基因编码的BUBR1蛋白则是纺锤体组装检查点(SAC)的核心成员,负责在细胞分裂时确保染色体正确分配,其变异可导致卵母细胞减数分裂中染色体分离错误,进而产生非整倍体胚胎。然而,PADI6复合杂合突变的具体组合形式多样、差异化分子致病机制尚待阐明;BUB1B单个拷贝(杂合)变异是否足以影响女性生殖,更一直缺乏直接的家系证据。北京家恩德运医院遗传中心的这两项研究,正是针对上述两个关键空白展开的系统性攻关。
成果一:PADI6"一截一延"复合杂合移码突变导致着床前胚胎致死
首报"一截一延"复合杂合移码突变组合
该研究纳入一对因PREMBL就诊于我院的夫妇。女方(先证者,33岁)有5年不孕史,曾接受两次IVF治疗(GnRH拮抗剂方案):第一周期取卵12枚、8枚受精,第二周期取卵15枚、9枚受精。然而,两次周期中所有卵裂期胚胎均在培养第3天(4~8细胞期)发生发育阻滞,无一进展至囊胚阶段,最终均未能移植。男方精液参数及激素水平均在正常范围内。
研究团队对先证者进行了系统性遗传学评估,包括染色体核型分析(550带G显带)、高分辨率染色体微阵列分析(CMA,Cytoscan 750k)及全外显子组测序(WES,Illumina NovaSeq 6000)。CMA结果显示基因组拷贝数正常,核型异常被判定为良性多态性。WES在PADI6基因(NM_207421.4)中检出两个双等位基因移码变异,经Sanger测序验证,分别来自父方和母方:
其一,c.707dupT(p.L237Afs*24)——来自父方的"截短型"突变:位于PADI6蛋白中间结构域,单碱基T的插入导致移码,引入提前终止密码子(PTC),产生仅含259个氨基酸的截短蛋白(较野生型694 aa缩短62.7%),C端结构域完全缺失。依据ACMG指南分类为"可能致病性"(PVS1+PM2+PP4)。
其二,c.2009_2010delAG(p.E670Gfs*48)——来自母方的"延长型"突变:位于PADI6蛋白C端结构域,2碱基AG的缺失导致移码,终止密码子延迟出现,产生含716个氨基酸的延长蛋白(较野生型多出22个氨基酸),C端局部结构发生显著改变。依据ACMG指南分类为"致病性"(PVS1+PM2+PP4)。
值得特别关注的是,这两个变异的复合杂合组合在PREMBL2患者中属首次报道,且在gnomAD等人群数据库中均未见收录,进一步支持其致病性。
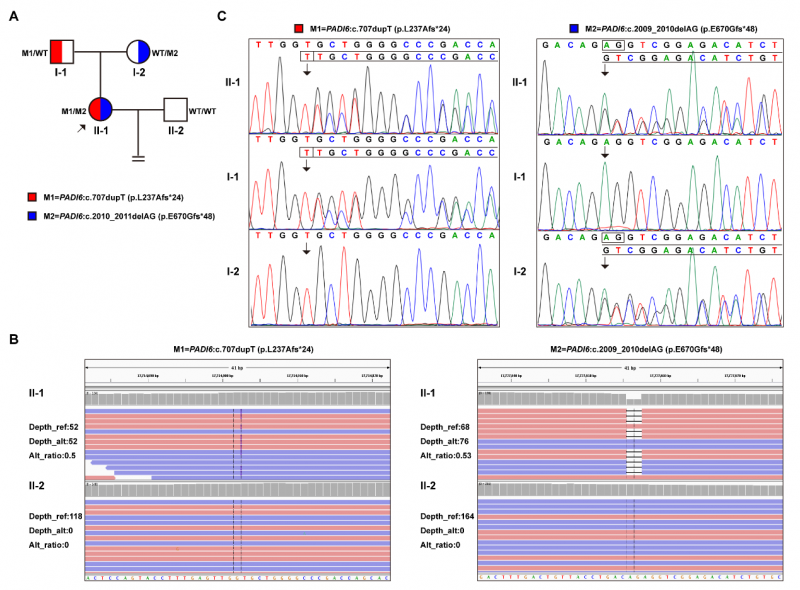
图1 全外显子组测序检测出两种PADI6移码突变
多维度解析差异化致病机制
本研究的突破性发现体现在以下四个层面:
1. 方向相反的双移码突变,揭示功能域差异化破坏。两个突变均为移码变异,但效应截然相反——c.707dupT导致蛋白截短(259 aa,缩短62.7%),c.2009_2010delAG导致蛋白延长(716 aa,延长3.2%)。这种"一截一延"的复合杂合模式在PADI6相关疾病中极为罕见,提示PADI6中间结构域与C端结构域在维持蛋白功能方面各有其不可替代的作用。
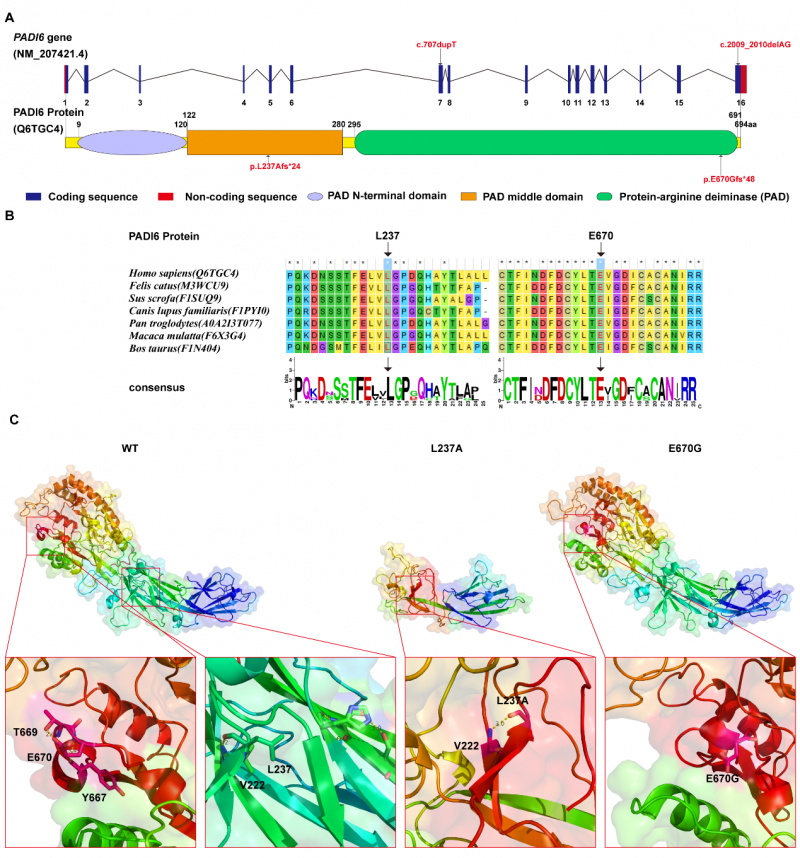
图2 变异的保守性分析及蛋白结构分析
2. 40 ns全原子分子动力学模拟,从原子层面阐明结构破坏机制。研究团队基于SWISS-MODEL同源建模,利用NAMD 2.9软件对野生型及两种突变体蛋白进行了40 ns全原子MD模拟。RMSD轨迹显示,L237A突变体整体构象极度不稳定;E670G突变体整体构象相对稳定,但C端在模拟后期出现明显局部漂移。RMSF分析进一步揭示:L237A突变体在第170位残基后α螺旋几乎完全消失、β折叠含量显著降低、无规卷曲大量增加;E670G突变体则在第660位残基后出现α螺旋去稳定化和β折叠减少。两种突变体在突变位点与邻近残基间的氢键数量均较野生型显著减少,从原子层面阐明了蛋白功能丧失的结构基础。
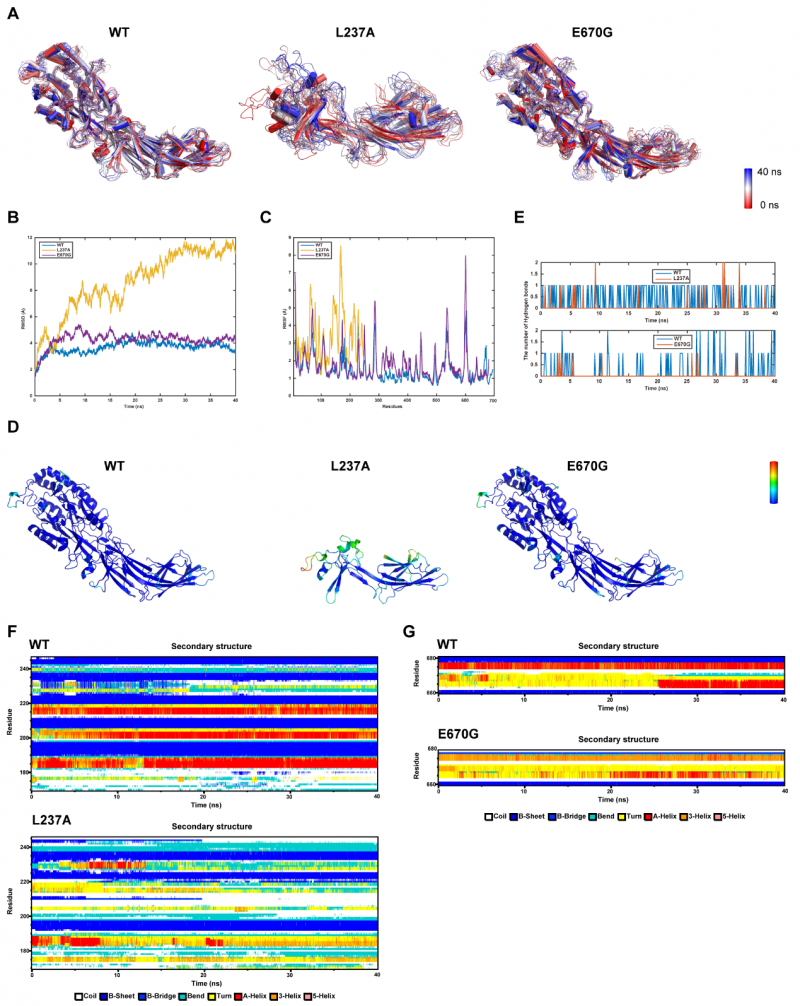
图3 40 ns全原子分子动力学模拟分析
3. 多层次细胞实验验证蛋白功能完全丧失。免疫荧光(IF)实验证实两种突变体亚细胞定位(胞质分布)与野生型一致,排除定位异常;但荧光强度均显著低于野生型。Western blot检测到L237A突变体条带位于约30 kDa(预期31 kDa),E670G突变体约80 kDa(预期82 kDa),两者蛋白表达量均显著下调。qPCR同时检测到PADI6及其相互作用蛋白(CPEB1、CFL1、BTF3、ZAR1、ANLN、ECT2、YBX2)的mRNA水平均显著降低,提示SCMC复合体的级联失稳效应。

图4 免疫荧光、qPCR和Western blot功能实验验证
4. 提出SCMC介导的分子致病模型,串联基因型与临床表型。基于上述多维度证据,研究团队提出了一个连贯的分子致病假说:PADI6功能丧失→皮质下母源复合体(SCMC)高级组装受阻→复合体弥散分布于胞质→无法整合入细胞质晶格→母源蛋白储存耗竭→母源-合子基因组转换(MZT)失败→胚胎在4~8细胞期发育阻滞。这一模型将基因变异、蛋白结构改变与临床表型有机串联,为PREMBL2的发病机制提供了系统性解释框架。
临床意义:从"反复失败"到"精准诊断"
对于反复IVF失败、胚胎均在卵裂期阻滞、且常规检查未见异常的女性不孕患者,应积极开展全外显子组测序或全基因组测序,重点关注PADI6等SCMC相关基因的复合杂合突变。明确遗传病因不仅有助于终结患者的"诊断漂流",更能为其提供精准的遗传咨询——包括再发风险评估(后代25%患病概率)、供卵辅助生殖的适应证判断,以及未来可能的基因治疗方向。
研究团队同时指出,本研究作为单病例报告,其结论尚需在更大规模的PREMBL患者队列中加以验证,并通过人类卵母细胞或动物模型的功能实验进一步确认因果关系。未来研究应聚焦于:在更多患者中筛查该复合杂合组合、开展mRNA稳定性及SCMC组装的直接功能验证,以及探索PADI6变异与其他生殖表型(如复发性流产、葡萄胎)之间的潜在关联。
成果二:BUB1B杂合变异导致复发性流产,揭示"父传女发"性别特异性
鉴定两种新型BUB1B变异,证实染色单体早熟分离表型
该研究对两个不相关的汉族复发性流产家系(家系1和家系2)进行了深入的遗传学分析。每个家系中的女性先证者均经历了3次或以上的不明原因流产。研究团队创新性地联合应用全外显子组测序(WES)、染色体微阵列分析(CMA)、高分辨染色体核型分析及着丝粒特异性PNA-FISH技术,从基因、染色体、蛋白等多个层面展开验证。
研究在家系1先证者中鉴定出c.2215G>T(p.A739S)变异,该变异遗传自临床表型正常的父亲,母亲和弟弟均未携带;在家系2先证者中鉴定出c.2164T>C(p.W722R)变异,先证者的姐姐(无流产史)及其两个儿子均未携带。两个变异均位于BUB1B基因第16号外显子、BUBR1蛋白高度保守的激酶结构域附近,在人群数据库(gnomAD)中极为罕见,ClinVar未见收录,多种生物信息学工具均预测为有害。这两种全球首次报道的BUB1B杂合错义变异,丰富了BUB1B相关疾病的突变谱,为理解杂合变异的生殖致病机制提供了新数据。
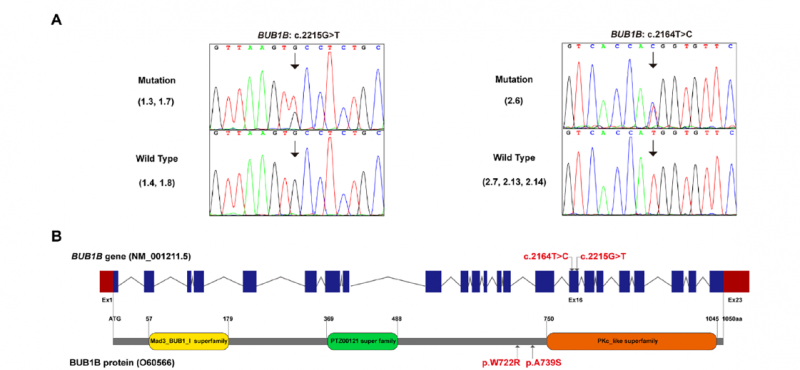
图5 BUB1B基因及BUBR1蛋白结构域示意图,箭头所指为两个错义变异(p.W722R、p.A739S)位置,均位于激酶结构域附近
通过高分辨染色体核型分析,两位先证者外周血淋巴细胞的染色单体早熟分离(PCS)率显著升高:家系1为8.93%(5/56),家系2为13.79%(8/58),而20名健康对照的平均PCS率仅0.40±1.05%(P<0.001)。进一步采用着丝粒特异性PNA-FISH精准定量,家系1的FISH-PCS率为18.58%(21/113),家系2为24.43%(32/131),对照平均仅2.95±0.60%(P<0.001)。对6份流产组织的CMA分析显示,每一份标本均呈现不同的随机非整倍体,提示非整倍体源于母体卵子的染色体分配缺陷,而非偶然的随机异常。
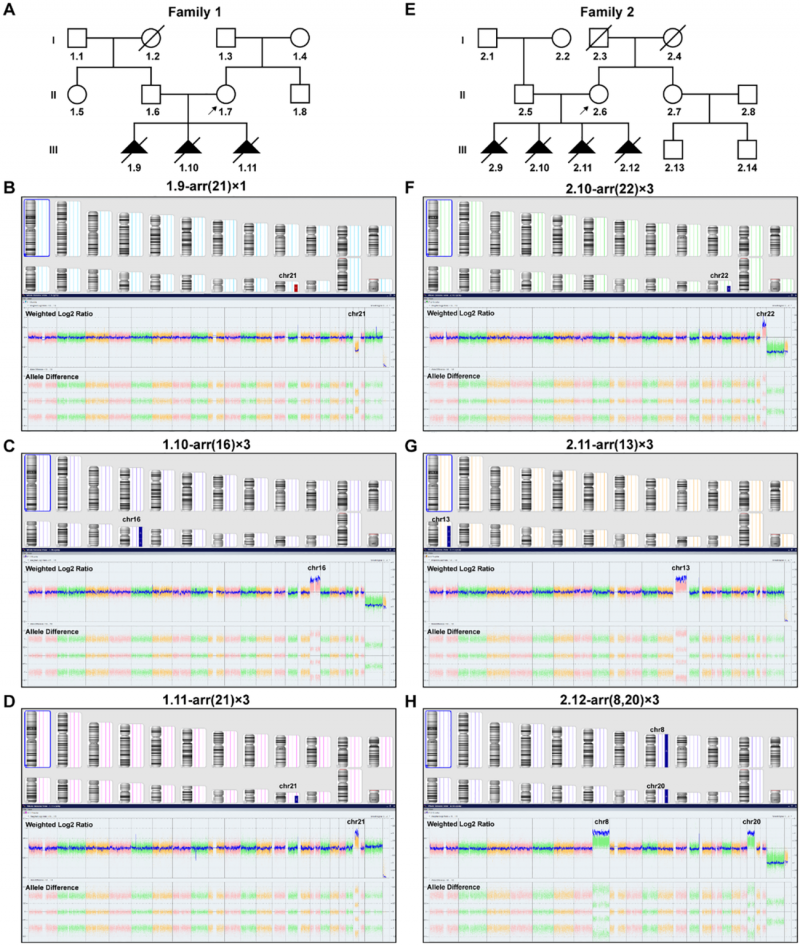
图6 两个家系的系谱图及流产组织染色体微阵列分析结果,每个流产标本均呈现不同的非整倍体核型
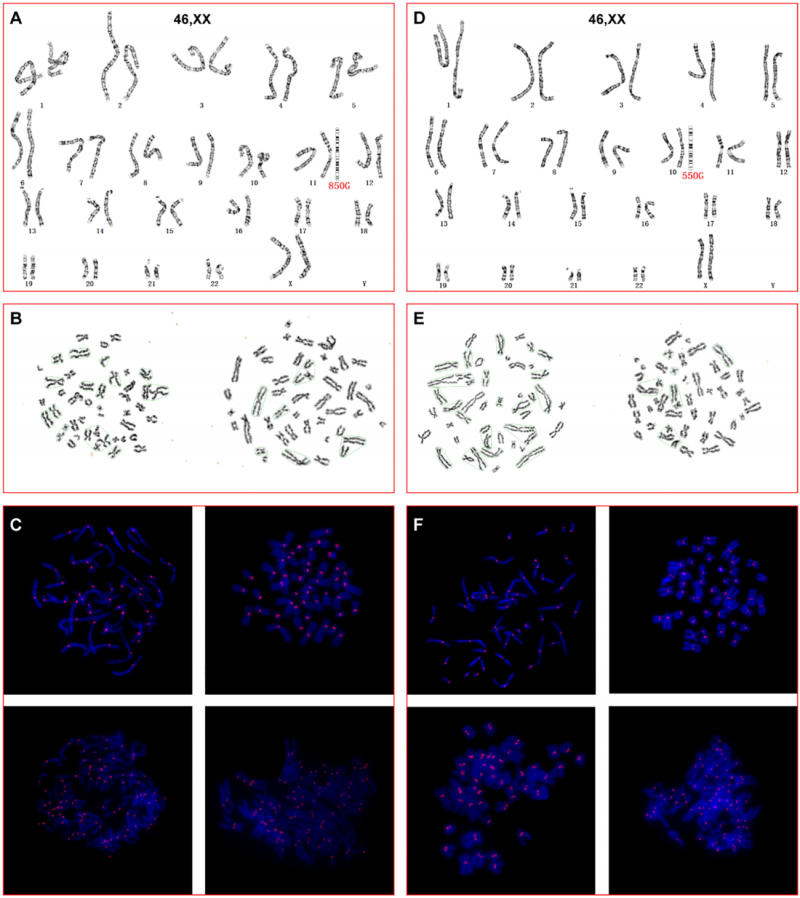
图7 先证者外周血淋巴细胞高分辨染色体核型及PNA-FISH分析,显示多条染色体出现染色单体早熟分离
阐明"父系传递、女性发病"的性别特异性外显机制
家系1中变异遗传自表型正常的父亲,这一现象的关键解释在于女性卵母细胞减数分裂对BUBR1蛋白的"高需求"。研究表明,BUBR1蛋白在卵母细胞从生发泡到MII期的成熟过程中表达量逐步升高,而卵母细胞的SAC本就比体细胞"宽松",无法对单个未附着的着丝粒做出响应。因此,即使BUBR1仅因杂合变异而部分减少,也可能足以突破卵母细胞本就脆弱的检查点,导致染色体提前分离、产生非整倍体卵子和胚胎。相比之下,男性生殖细胞对SAC蛋白剂量不敏感,这解释了父亲作为携带者生育正常、而女儿携带变异后反复流产的现象。
在功能层面,蛋白结构模拟显示,p.W722R变异以带正电的精氨酸取代疏水性色氨酸,破坏局部疏水核心,可能影响蛋白稳定性;p.A739S变异以极性丝氨酸取代非极性丙氨酸,可能干扰BUBR1与下游效应分子的相互作用。RT-PCR和Western blot检测显示,两位先证者的BUB1B mRNA和BUBR1蛋白表达均呈下降趋势,家系1的父亲(杂合携带者)也有类似趋势,提示"单倍剂量不足(haploinsufficiency)"致病机制。
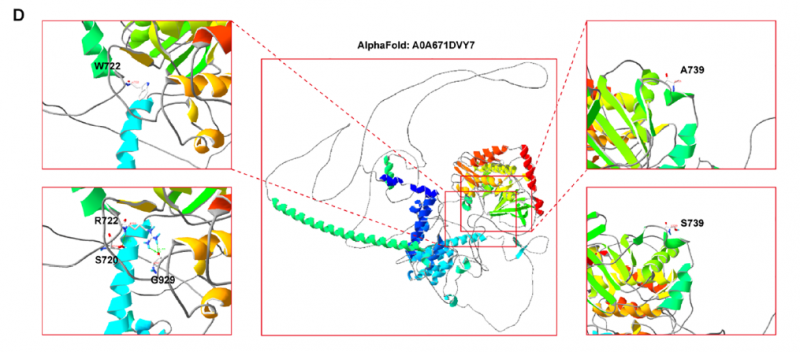
图8 错义变异的BUBR1激酶结构域三维结构模拟,显示变异导致蛋白构象改变
临床意义:从"查不出原因"迈向"精准遗传咨询"
此项研究成果对临床实践具有重要的指导价值:
其一,对于临床表现为反复流产且外周血检出PCS表型的女性,建议将BUB1B纳入靶向基因筛查,而非对所有RPL患者常规筛查,以提高检测的针对性与性价比。
其二,携带BUB1B变异的RPL女性,建议行胚胎植入前非整倍体遗传学检测(PGT-A),筛选整倍体胚胎移植,提高活产率、降低流产风险。
其三,鉴于BUB1B变异与早发性卵巢功能不全(POI)的关联,若反复无法获得可移植的整倍体胚胎或卵巢储备明显下降,可考虑供卵IVF。
其四,遗传咨询时需特别注意"父系传递、女性发病"的性别特异性外显特点——BUB1B变异可由临床表型正常的男性携带者传递,这一信息对家系风险评估至关重要。
双研究联动:覆盖"着床前—着床后"全链条的生殖遗传诊断体系
将两项研究并置审视,可见一条清晰的科学逻辑主线:女性生殖失败可发生于胚胎发育的不同阶段,而其背后往往是不同基因、不同机制导致的卵母细胞或早期胚胎功能缺陷。PADI6突变破坏的是卵母细胞的"蛋白储备仓库"——细胞质晶格与SCMC复合体,使胚胎在母源-合子基因组转换之前便因母源因子耗竭而停滞于4~8细胞期,表现为"无胚胎可移植";BUB1B杂合变异破坏的则是卵母细胞减数分裂的"染色体质检关卡"——纺锤体组装检查点,使染色体分离错误得以"漏网",产生非整倍体卵子与胚胎,表现为"能着床却反复流产"。前者阻断在着床之前,后者失败在着床之后,两者恰好覆盖了女性生殖障碍的关键时间窗。
从临床转化角度看,两项研究共同指向一个核心理念:对于反复生殖失败的患者,应依据其临床表型精准选择遗传检测策略。胚胎卵裂期阻滞者,优先排查PADI6等SCMC相关基因的复合杂合突变;反复流产且检出PCS表型者,则将BUB1B纳入靶向筛查。明确遗传病因后,方可为患者提供从再发风险评估、PGT胚胎筛选到供卵IVF的阶梯式精准生育指导,真正帮助患者走出"反复失败—反复检查—病因不明"的恶性循环。
团队实力:深耕生殖遗传,守护生育健康
北京家恩德运医院医学遗传中心始终坚持"临床驱动科研,科研反哺临床"的理念。继2025年在《Human Genomics》《Clinica Chimica Acta》刊发先天性并指畸形、遗传性鱼鳞病相关研究,以及2026年初遗传性鱼鳞病(FLG、ABCA12基因研究)方向发表SCI论文后,PADI6与BUB1B这两项成果进一步彰显了团队在罕见生殖遗传病领域的深厚积累。PADI6研究由遗传中心检验师焦顺焘为第一作者,生殖中心谭姿辉医生、遗传中心魏天颖主任为共同第一作者,胡华莹主任、刘家恩教授为共同通讯作者;BUB1B研究由魏天颖主任担任第一作者,胡华莹主任担任通讯作者。
随着全外显子组测序成本的降低和各种遗传检测技术的广泛交叉应用,针对女性生殖障碍的精准遗传诊断正变得日益可行。北京家恩德运医院将继续深耕基因组学领域,致力于将前沿科研成果转化为临床实践,为更多深受不孕与反复流产困扰的家庭孕育新希望。
分享到微博



